GCRA
OVERVIEW
FAQ
NEWS
ARTICLES
PHOTOS
REEF ISSUES
RESTORATION
PAPERS
LINKS
|
|
|
|
Increased Zooxanthellae Numbers and Mitotic Symbiosis (2004) 37:107-120 THOMAS J. GOREAU1*, JAMES M. CERVINO2, and RACHAEL POLLINA3 1Global Coral Reef Alliance, 37 Pleasant Street, Cambridge, MA 02139, Tel. +1-617-864-4226, Email. goreau@bestweb.net; 2Department of Marine Science, University of South Carolina, Columbia, SC 29073, Tel. +1-917-620-5287, Email. cnidaria@earhlink.net; 3Department of Microbiology, University of Massachusetts, Amherst, MA 01003, USA, Tel. +1-413-577-2939, Email. Cellulah@aol.com Received December 1, 2003; Accepted March 15, 2004Abstract Zooxanthella densities, mitotic indices, and chlorophyll were measured in six major reef-building coral genera growing on electrically stimulated "Biorock" reefs in Indonesia, and compared with genetically identical corals growing adjacent to them. Corals on Biorock reefs had generally higher densities of zooxanthellae and higher rates of symbiotic algal division, but generally lower chlorophyll per zooxanthella. These patterns are coincident with higher coral skeletal growth rates, better developed branching morphology, and higher rates of resistance to environmental stresses, including elevated temperatures, nutrients, and sediments. Zooxanthellae play a critical role in providing alkalinity for coral calcification by withdrawing carbon dioxide for photosynthesis, and energy for proton and calcium pumping. Our results support the view that Biorock corals are healthier than normal corals because alkalinity for calcification is provided by elevated pH caused by electrolysis of seawater. Presented at the 4th International Symbiosis Congress, August 17-23, 2003, Halifax, Canada *The author to whom correspondence should be sent. 0334-5114/2004/$05.50 ©2004 Balaban By providing alkalinity for skeletal growth directly, the Biorock process reduces the large energy sink of proton and calcium pumping for skeletal formation, leaving the coral more metabolic energy for growth, reproduction, and resisting environmental stresses. Because Biorock corals have the symbiotic coralzooxanthella relationship de-coupled under non-stressed conditions, they provide a new experimental tool for examining regulation of zooxanthella densities. Because the effects of increased coral calcification on zooxanthellae was small, corals appear to be more dependent on their algae than the converse. Since the Biorock process is unique in greatly increasing coral survival from bleaching and pollution, and in reversing the increasing acidity caused by rising global carbon dioxide, it may prove crucial to the long term survival of coral reef species and ecosystem function. Keywords: Zooxanthella, coral, Mitotic Index, electrical stimulation, Biorock process, symbiosis regulation, stress resistance, calcification
1. Introduction There is a critical need to develop methods capable of restoring coral growth and the ecosystem services that coral reefs provide. Corals have long been known to be very vulnerable to small environmental changes (Goreau et al., 1979). The last few decades have shown just how sensitive they are by rapidly increasing death of corals, primarily from heat stress, diseases, and algae overgrowth, as well as sediments, dynamite and bombs, poisons, dredging, and boat and diving activities. Most habitats that are now classified as "coral" reefs are in fact coral communities, in which hard corals form an increasingly small portion. As a result almost none of the reefs which were documented in the world's largest collection of coral reef images from the 1940s, 1950s, and 1960s to be almost totally covered by hard corals now have more than a small portion of their former abundance (T.J. Goreau, in preparation). There is now very little pristine reef left to protect, and vast areas are severely degraded, with resulting loss of their biodiversity, fisheries production, ecotourism value, and shore protection services. Restoring these degraded areas is critical for the coastal zone economy of over 100 countries. Restoration in the face of globally increasing temperatures, disease, weedy algae growth, and sedimentation requires approaches that enhance coral growth so they can tolerate greater stresses than existing coral populations can. The only method known that does so is the "Biorock" method (Hilbertz and Goreau, 1996). Biorock projects in more than a dozen countries around the world have shown extremely rapid growth of corals, up to several times greater than normal, even in habitats where all other corals, including controls, were killed by algae overgrowth (Goreau and Hilbertz, 1996, Fig. 1a). Corals grown on ZOOXANTHELLAE DENSITY AND DIVISION 109 Biorock coral nurseries in the sea bleached like coral in surrounding reefs, but had 16 to 50 times greater survival from bleaching following the 1998 Bleaching Event in the Maldives (Goreau et al., 2000a,b: Fig. 1b,c). Hundreds of coral transplants had been made in adjacent sites by conventional cementing or epoxy methods. All of the controls died after bleaching. Normally the alkalinity required for coral skeletal deposition of aragonitic calcium carbonate is provided by the removal of carbon dioxide by zooxanthella photosynthesis, which pulls the bicarbonate system equilibrium to the right:
2HC03 = H2O + CO2 + C03=
The resulting alkalinity is neutralized by precipitation of calcium carbonate, with a near one to one ratio between photosynthesis and calcification. Because of the physical separation of photosynthesis and calcification, the process requires active transport of calcium and protons (Goreau and Goreau, 1959). It is not clear what effect decoupling calcification from photosynthesis will have on the symbiotic algae, and the Biorock process provides an experimental tool for examining this interaction. The basis for increased growth and stress tolerance of Biorock corals is thought to be because the coral benefits from elevated pH at the surface of the growing skeleton provided by low voltage seawater electrolysis. At the cathode alkalinity is generated through the electrochemical hydrolysis of water:
2e + 2H20 = H2 + 201-1
As a result of this alkalinity generation, one hydroxyl ion is produced per electron. This causes the electrolytic precipitation of aragonite, mixed with magnesium hydroxide from seawater. Although ocean surface waters are thermodynamically supersaturated n calcium carbonate, precipitation does not happen because of kinetic barriers to nucleation, which are overcome by the electrical current. It seems that the availability of alkalinity at the surface of the coral skeleton increases skeleton growth, so less coral metabolic energy is required for proton and calcium pumping, leaving extra energy for growth, reproduction, and resisting environmental stresses. Corals grown on Biorock have visibly higher coral skeletal growth rates, often 3-5 times normal (Goreau and Hilbertz, 1996; P.N. Dwija, in preparation), brighter tissue color, increased polyp extension, and better developed branching morphology (Figs. la-c). Branching and foliose species tend to elongate rapidly perpendicular to the Biorock substrate while encrusting species grow rapidly 110 T.J. GOREAU ET AL. along the substrate. They attract dense populations of adult and juvenile resident and migratory fishes (Goreau et al., 2000a). Despite successful field results from nearly a hundred projects in over a dozen countries around the world, there has been little or no effective support for large-scale restoration of reefs or efforts to set up nurseries of more resistant corals. Part of this resistance has been based on claims that there was no "evidence" other than direct field observations and images that the corals benefiting from the Biorock process were different from normal corals (other than surviving normally lethal stresses). In this paper we examine the metabolic health of Biorock corals by comparing the zooxanthellae density, mitotic indices, chlorophyll densities, and chlorophyll per zooxanthella of six genera of reef building corals with genetically identical controls. Results of coral growth rate measurements, coral skeletal chemistry, and histological analysis of tissue reproductive status will be published separately (P.N. Dwija, in preparation; Goreau et al., in preparation).
2. Materials and Methods Small portions of corals of six genera (Acropora nasuta, Pocillopora verrueosa, Montipora capitata, Seriatopora hystrix, Porites cylindrica, and Turbinaria renifromis), typically of the order of ten centimeters across, were transplanted from corals growing in the coral reef of the Pemuteran Marine Protected Area, Bali, Indonesia onto lower portions of Biorock coral nurseries a few meters away. After a year of growth, 4-8 sub-samples of small growing tips, a few centimeters in length, were taken from the Biorock colonies along with a similar number of similarly sized portions of the control mother colony. Because they are genetically identical and growing in the same physical habitat the only difference should be the effects of the electrical currents. Samples were preserved in 10% gluteraldehyde in seawater. In the laboratory a sub-sample was taken for chlorophyll analysis. This subsample had the tissue re-suspended in seawater by rubbing with a stiff brush. The suspension and solution was then extracted with acetone for standard spectrophotometric analysis of chlorophyll a and c2 peaks at 630 nm and 750 nm See figure on next page. Figure 1. a-c: Corals on Pemuteran Biorock reefs from which samples were taken, showing healthy growth. All photographs by Wolf Hilbertz. a. Encrusting coral growing along Biorock substrate. b. Coral showing rapid growth and dense new branch formation. c. Dense branching of coral and fish attracted to it.
ZOOXANTHELLAE DENSITY AND DIVISION 111
112 T.J. GOREAU ET AL. respectively (Jeffrey and Humphrey, 1976). The rest of the sample was used for analysis of zooxanthellae. The tissue was water-picked and centrifuged at 5,000 rpm for 5 minutes. The resulting zooxanthella pellet was re-suspended in 10 milliliters of seawater and a 100 microliter sub-sample placed on a reticular grid microscope slide. Ten replicates were used for each sample. Total numbers of zooxanthellae were counted, along with the number of cell doublets (Wilkerson et al., 1988; Suharsono and Brown, 1992). The number of doublets, divided by the total number of cells, gives the mitotic index in percent. Zooxanthella densities and chlorophyll absorbances were ratioed to the weight of the coral samples. Chlorophyll density divided by zooxanthella density gave the chlorophyll per zooxanthella. As chlorophyll measurements were the absorbance of the total sample analyzed, no statistics could be computed for these values.
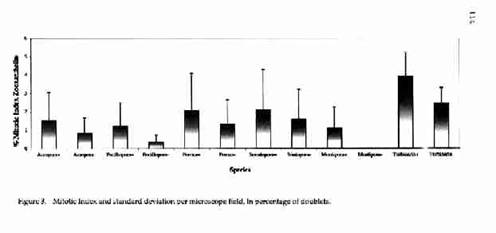
3. Results The results are shown in Figs. 2-5, representing the zooxanthella densities, the mitotic indices, the chlorophyll densities, and the chlorophyll per zooxanthella respectively of all Biorock corals and controls. Five of the six genera had markedly higher zooxanthella densities in Biorock corals than controls (Fig. 2). Only Porites showed slightly lower values in Biorock corals. The difference was very highly statistically significant at for Seriatopora and for Turbinaria (Table 1).
NS-Not significant, *Significant at 0.05 level, **Highly significant at 0.01 level, ***Very highly significant at 0.001 level.
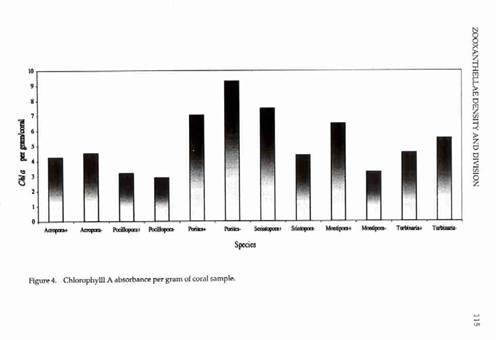
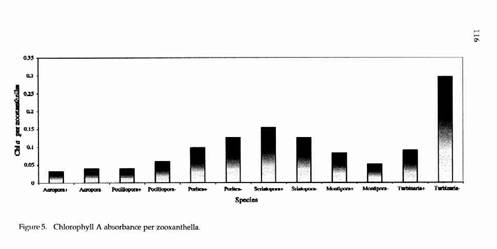
ZOOXANTHELLAE DENSITY AND DIVISION 117 All six genera had higher mitotic indices in Biorock corals than controls (Fig. 3). The mitotic index of Montipora controls was almost zero. The difference was statistically significant for Acropora, highly significant for Pocillopora, and very highly significant for Montipora (Table 1). Chlorophyll densities were higher in Biorock corals than controls in three genera (Pocillopora, Seriatopora, and Montipora) and lower in three (Acropora, Porites, and Turbinaria) (Fig. 4). Chlorophyll densities per zooxanthella were higher in two genera (Seriatopora and Montipora) and lower in four (Acropora, Pocillopora, Porites, and Turbinaria) (Fig. 5). Microscopic examination of all specimens showed no visible abnormalities in the tissue or skeleton of Biorock corals compared to controls. However, Biorock corals do show a denser and more fully developed macroscopic branching pattern.
4. Discussion These generally higher zooxanthella densities and mitotic indices are results are consistent with Biorock corals having improved growth compared to controls, as is observed for skeletal growth, and increased resistance to stress. However, due to small sample sizes in this preliminary study, more replicates are needed to reduce sample variances and improve statistical confidence limits. High zooxanthella densities and high mitotic indices are signs of a healthy coral whose symbiotic algae are rapidly proliferating (Belda et al., 1993; Muller-Parker et al., 1994; Stambler et al., 1994). This suggests that zooxanthellae are more efficient at growth under Biorock conditions. All species showed higher mitotic indices and 5 of 6 had higher zooxanthella densities in Biorock corals compared to controls. In stressed corals high mitotic indices also occur, but this is found under low zooxanthella densities and is thought to be a stress response in which the zooxanthella division rates rise to compensate for their loss (Jones, 1997; Jones and Yellowlees, 1997). Lowered chlorophyll content in half the genera, and in particular lower chlorophyll per zooxanthella in two thirds of the genera, of Biorock corals compared to controls may be consistent with rapid growth if it is a result of cellular mechanisms to regulate "excessive" proliferation under rapid growth conditions. It could be analogous to the lower chlorophyll content of sun adapted corals compared to shade adapted species of corals (Falkowski and Dubinsky, 1981). This is thought to be a mechanism to reduce photosynthesis rates under conditions of high growth rate, and possibly reduce photooxidative stress. The lowered chlorophyll could result from a cellular regulatory mechanism to reduce the rates of possibly deleterious side products 118 T.J. GOREAU ET AL. of photosynthesis, such as excessive alkalinity generation (Kuhl et al., 1995) resulting from carbon dioxide removal and fixation, or uncontrolled electron flows (Iglesias-Prieto et al., 1992; Warner et al., 1999). The differences found in .chlorophyll per zoxanthella in different genera in response to the Biorock process suggests that different physiological regulatory pathways are operating. Coral calcification is normally closely coupled to zooxanthella photosynthesis, which is thought to result from the increased alkalinity following carbon dioxide fixation, linked to active proton and calcium pumping (Goreau and Goreau, 1959). However, if alkalinity is de-coupled from photosynthesis, as it is under Biorock conditions by the direct elevation of pH resulting from electrolysis of water, then the near one to one ratio of carbon flows into photosynthesis and calcification could be disrupted. Coupling of proton and calcium pumping (Goreau and Goreau, 1959; Al Horani et al., 2003) could be disrupted if there is less need for this large metabolic expenditure due to the coral getting the conditions for rapid calcification for free from the electrolytic Biorock process. The modest effect of increased pH and calcification on zooxanthella densities suggests that increased calcification does not markedly stimulate photosynthesis, as has been speculated McConnaughey and Whelan, 1996). This suggests that under normal growth conditions corals are more dependent on their zooxanthellae than the converse. Experimental conditions previously studied that disrupt symbiosis, such as extreme temperatures, salinities, chemicals, or diseases, generally stress one or both symbionts. In contrast, the decoupling under Biorock conditions appears to be under conditions of good health and high growth of both partners. New insights into the regulation of calcification, photosynthesis, pH homeostasis, active transport, zooxanthella density, carbon flow pathways, and metabolic regulation of trace metal and stable isotope composition of coral skeleton could therefore be gained by application of the Biorock method under different conditions. Further insights would result if these measurements are coupled to microelectrode analysis of pH, calcium ion activity, oxygen, and carbon dioxide concentrations, PAM fluorometry, enzyme assays, skeletal trace metal and stable isotope chemistry, and assays of genetic expression in host and symbiont. The Biorock process is unique in greatly increasing coral survival from bleaching and pollution, and in locally reversing the increasing acidity caused by rising global carbon dioxide. It may therefore prove crucial to the long-term survival of coral reef species and ecosystem function as globally increasing temperature and pollution kill corals at increasing rates.
Acknowledgments We thank Wolf Hilbertz and Randall Dodge for assistance in the field, and Taman Sari Hotel for supporting the project. We are grateful to support from the Leslie Jones Foundation that supported the field lab work in this project. We thank Ray Hayes for help with statistical calculations and Bob Trench for discussions. REFERENCES AI-Horani, F.A., Al-Mograbi, S.M., and De Beer, D. 2003. The mechanism of calcification and its relationship to photosynthesis and respiration in the scleractinian coral Galaxea fascicularis. Marine Biology 142: 419-426. Belda, C.A., Lucas, J.S., and Yellowlees, D. 1993. Nutrient limitation in the giant clamzooxanthellae symbiosis: effects of nutrient supplements on growth of the symbiotic partners. Marine Biology 117: 655-664. Falkowski, P.G. and Dubinsky, Z. 1981. Light -shade adaptation of Stylophora pistillata, a hermatypic coral from the Gulf of Elat. Nature 289: 172-174. Goreau, T.F. and Goreau, N.I. 1959. The physiology of skeleton formation in corals. II. Calcium deposition by hermatypic corals under various conditions in the reef. Annals New York Zoological Society 239-250. Goreau, T.F., Goreau, N.I., and Goreau, T.J. 1979. Corals and coral reefs. Scientific American 241: 124-136. Goreau, T.J. and Hilbertz, W. 1996. Reef restoration using seawater electrolysis in Jamaica, http:/ /www.globalcoral.org/ reef restoration using_seawater.htm. Goreau, T.J., Hilbertz, W., and Azeez A. Hakeem, A. 2000. Increased coral and fish survival on mineral accretion reef structures in the Maldives after the 1998 Bleaching Event, http:/ /www.globalcoral.org/Increased%2OCoral%20and%2OFish%2OSurvival%2Oo n%20Mineral%20Accretion.htm. Goreau, T.J., Hilbertz, W., Azeez A. Hakeem, A., and Hameed, S. 2000. Reef restoration and shore protection projects at Ihuru tourist resort, Republic of Maldives, using mineral accretion: preliminary results, http: / /www.globalcoral.org/Reef%20Restoration%2OShore%2OProtection%2OAt%2 Olhuru%20%20Maldives,htm. Hilbertz, W.H. and Goreau, T.J. 1996. Method of enhancing the growth of aquatic organisms, and structures created thereby, United States Patent Number 5,543,034, U.S. Patent Office (14 pp). Iglesias-Prieto, R., Roberto, J., Matta, L., Robins, W.A., and Trench, R.K. 1992. Photosynthetic response to elevated temperature in the symbiotic dinoflagellate Symbiodinium microadriaticum in culture. Proceedings of the National Academy of Sciences USA 89: 10302-10305. Jeffery, S.W. and Humphrey, G.F. 1976. New spectrophotometric equations for determining Chl a,b,c & C2 in higher plants, algae and natural phytoplankton. Biochemie and Physiologie der Pflanzen 167: 191-194 . Jones, R.J. 1997. Zooxanthellae loss as a bioassay for assessing stress in corals. Marine Ecology Progress Series 149: 163- 171. Jones, R.J. and Yellowlees, D. 1997. Regulation and control of intracellular algae (=zooxanthellae) in hard corals. Philosophical Transactions Royal Society London B 352: 457-468. Kuhl, M., Cohen, Y., Dalsgaard, T., Jorgensen, B.B., and Revsbech, N.P. 1995. Microenvironment and photosynthesis of zooxanthellae in scleractinian corals studied with microsensors for 02, pH and light. Marine Ecology Progress Series 117: 159-172. McConnaughey, T.A. and Whelan, J.F. 1996. Calcification generates protons for' nutrient and bicarbonate uptake. Earth Science Reviews 967: 1-23. Muller-Parker, G., McCloskey, L.R., Hoegh-Guldberg, 0., and McAuley, P.J. 1994. Effect of ammonium enrichment on animal and algal biomass of the coral Pocillopora damicornis. Pacific Science 48: 273-283. Stambler, N., Cox, E.F., and Vago, R. 1994. Effect of ammonium enrichment on respiration, zooxanthellae densities, and pigment concentrations in two species of Hawaiian corals. Pacific Science 48: 284-290. Suharsono, R.K. and Brown, B.E. 1992. Comparative measurements of mitotic index in zooxanthellae from a symbiotic cnidarian subject to temperature increase. Journal Experimental Marine Biology Ecology 158: 179-188. Warner, M.E., Fitt, W.K., and Schmidt, G.W. 1999. Damage to photosystem II in symbiotic dinoflagellates: a determinant of coral bleaching. Proceedings of the National Academy of Sciences USA 96: 8007-8012. Wilkerson, F.P., Kobayashi, D., and Muscatine, L. 1988. Mitotic Index of symbiotic algae in Caribbean reef corals. Coral Reefs 7: 29-36. |
| ||||||||||||||||||||||||||||||||||||||||||||||